Imunobiológia
Obsah
Imunoglobulíny
Už na konci 19. storočia bolo známe, že krvné sérum človeka a živočíchov, ktoré prekonali určité infekčné ochorenie, obsahuje nejaké látky schopné ochrániť týchto jedincov pred opakovaným tým istým ochorením. Takéto sérum dokázalo zhlukovať (aglutinovať) mikróbnych pôvodcov ochorenia, vyzrážať (precipitovať) rozpustné mikróbne antigény alebo neutralizovať (zneškodňovať) baktériové toxíny. Účinné látky v sére sa podľa týchto vlastností nazývali aglutiníny, precipitíny alebo antitoxíny. V roku 1890 ich Paul Ehrlich označil ako protilátky a tento názov sa rýchlo rozšíril. Vychádzal z toho, že protilátky sú látky pôsobiace proti telám mikróbov, ktoré vnikli do organizmu. V roku 1930 sa dokázalo, že protilátky sú bielkovinovej povahy a nachádzajú sa v gamaglobulínovej frakcii séra; odtiaľ pochádza aj ich dnes ešte občas používaný názov gamaglobulíny. Neskôr sa zistilo, že protilátková aktivita sa nachádza aj v betaglobulínovej frakcii, a preto po spoznaní ich základnej štruktúry sa nomenklatúrna komisia Svetovej zdravotníckej organizácie v roku 1964 uzniesla označiť protilátky názvom imunoglobulíny (skratka Ig).
Imunoglobulíny sú základnými molekulami imunitného systému. Z chemického hľadiska ide o glykoproteíny, ktorých prvoradou funkciou je protilátková aktivita. Nachádzajú sa v membráne na povrchu B lymfocytov, kde pôsobia ako receptor pre antigén. Rovnakú chemickú podstatu majú aj protilátky - sú to v podstate rozpustné (sekretované) formy B- lymfocytových receptorov.
{kind=link}
Štruktúra imunoglobulínov
{kind=link}
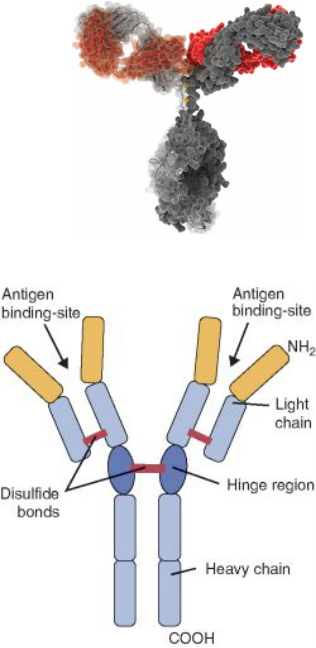
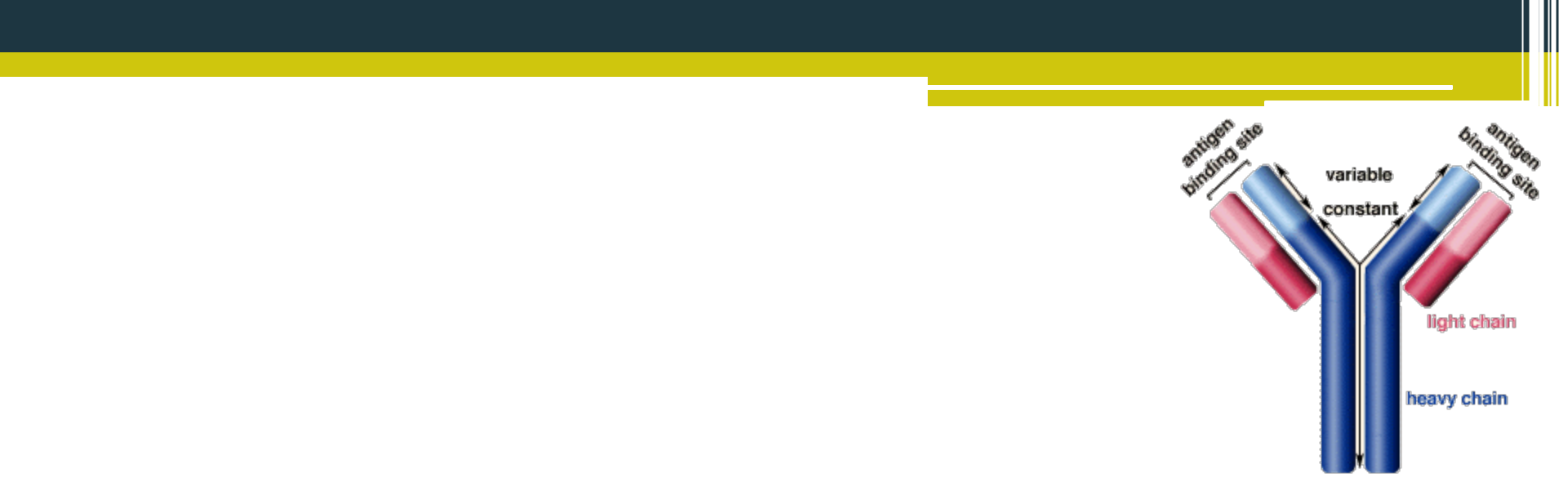
Základná molekula imunoglobulínu je zložená zo štyroch polypeptidových reťazcov : dvoch ťažkých (H - heavy) s rovnakou primárnou štruktúrou a dvoch ľahkých (L - light), ktoré tiež majú rovnakú primárnu štruktúru. Ťažké reťazce obsahujú približne 445 aminokyselín, ľahké reťazce pozostávajú z asi 214 aminokyselín. Reťazce sú medzi sebou pospájané disulfidovými väzbami. Každý reťazec (H aj L) pozostáva z dvoch základných oblastí: konštantnej oblasti (C - constant), t. j. úseku s relatívne nemennou sekvenciou aminokyselín pri všetkých imunoglobulínoch príslušného typu H alebo L reťazca a z variabilnej oblasti (V - variable), t. j. úseku, v ktorom sa imunoglobulíny príslušného typu H alebo L reťazca navzájom líšia. Konštantnú oblasť ťažkého reťazca označujeme CH, ľahkého CL; variabilná oblasť ťažkého reťazca sa označuje VH, ľahkého reťazca VL. Variabilné oblasti obsahujú aj tri hypervariabilné úseky, ktoré sa vyznačujú zvlášť vysokou variabilitou v primárnej štruktúre a tvoria väzbové miesta pre antigén. Špecifita imunoglobulínu vzhľadom k antigénu je podmienená predovšetkým primárnou štruktúrou hypervariabilných úsekov. Imunoglobulíny sú na rôznych miestach H reťazcov glykozylované - pripájajú sa na ne sacharidy N- glykozidovou väzbou. Ľahké reťazce sa rozdeľujú na dva typy: K (kappa) a X (lambda). Imunoglobulíny s rovnakým typom ľahkých reťazcov majú rovnakú konštantnú oblasť a navzájom sa líšia vo variabilnej oblasti. Variabilné a konštantné oblasti ľahkých reťazcov sa označujú VK, Vx, CK, Cx, pričom VK sa v reťazci vždy spája s CK, kým Vx sa spája s Cx. U človeka existuje päť základných typov ťažkých reťazcov: Y (gama), a (alfa), p (mí), ô (delta), E (epsilon). Imunoglobulíny s rovnakým typom ťažkých reťazcov majú rovnakú konštantnú oblasť a navzájom sa líšia vo variabilnej oblasti.
{kind=link}
Pôsobením proteolytických enzýmov na molekulu imunoglobulínu sa získajú charakteristické fragmenty. Papaín štiepi molekulu nad disulfidovými väzbami, ktoré spájajú oba H reťazce, pričom vznikajú dva identické fragmenty označované skratkou Fab a jeden fragment, ktorý sa označuje Fc. Názov Fab vyplýva z jeho schopnosti viazať antigén (fragment antigen binding) a skladá sa z L reťazca a časti H reťazca. Fragment Fc ľahko kryštalizuje, z čoho sa odvodzuje aj jeho názov (fragment crystalizable). Predstavuje koncovú časť imunoglobulínovej molekuly, čiže sa skladá zo zvyškových častí H reťazcov, spojených disulfidovými väzbami. Zodpovedá za biologické vlastnosti protilátky (väzba komplementu, opsonizácia, väzba k bunkovým receptorom). Pôsobením pepsínu alebo trypsínu vzniká fragment F. ). Je to podmienené tým, že štiepne miesto uvedených enzýmov je pod miestom disulfidových väzieb spájajúcich H reťazce, ktoré držia oba Fab fragmenty spolu. Fragment Fíab'h je takto bivalentný (obsahuje obidve väzbové miesta pre antigénový determinant), na rozdiel od fragmentov Fab, ktoré sú monovalentné (obsahujú len jedno väzbové miesto pre antigénový determinant).
{kind=link}
Reťazec H aj L sa delí na domény, pričom každá z nich sa skladá z približne 110 aminokyselinových jednotiek. Ľahký reťazec má dve domény (VL, CL). VL predstavuje variabilnú oblasť, ktorá sa podieľa na väzbe antigénu; CL predstavuje konštantnú oblasť, rovnakú pri všetkých protilátkach. Ťažký reťazec má jednu doménu VH a viacero domén CH - CH1, CH2, CH3, pričom pri Ig M a Ig E pribúda ešte doména CH4. Každá doména má vlastnú intradoménovú disulfidovú väzbu. Medzi doménami CH1 a CH2, s výnimkou molekúl Ig M a Ig E, sa nachádza pántová oblasť, ktorá je flexibilná a umožňuje voľnosť pohybu pre fragmenty Fab. Pántovú oblasť tvoria aminokyselinové jednotky v počte od približne 10 (pri reťazcoch al, a2, yl, Y2 a Y4) až vyše 60 (pri reťazcoch Y3 a ô).
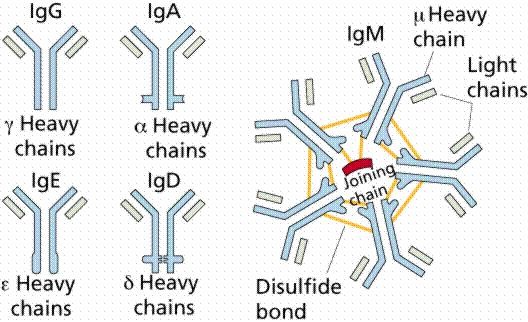
Rozdelenie imunoglobulínov
Imunoglobulíny jednotlivých tried sa odlišujú biologickou funkciou a výskytom. U človeka rozlišujeme 5 základných tried imunoglobulínov:
- Ig G tvorí až 80 % všetkých imunoglobulínov v krvnom sére. Tvorí sa najmä po opakovanej imunizácii (po druhom a ďalšom kontakte s antigénom) a po imunizácii rozpustným antigénom. Významne sa zúčastňuje opsonizácie, pričom konštantné oblasti H reťazcov (Fc oblasti) sú rozpoznávané fagocytmi. Týmto spôsobom fagocyty rozpoznajú a fagocytujú opsonizovaný objekt. Uplatňuje sa aj pri tvorbe imunokomplexov, aktivuje komplement klasickou cestou aktivácie. U primátov, hlodavcov a šeliem prestupuje placentou a zaisťuje obranu plodu. Je hlavným imunoglobulínom kolostra (prvé mlieko produkované po pôrode) najmä u druhov, u ktorých neprechádza placentou (u ošípanej, prežúvavcov, koní). U niektorých živočíšnych druhov majú imunoglobulíny funkčne a kvantitatívne zodpovedajúce Ig G natoľko odlišnú stavbu a vlastnosti, že sa pre ne používa zvláštne označenie (u kury domácej Ig Y, u koňa Ig T).
- Ig M sa vyskytuje v dvoch formách: ako monomér a pentamér. Monomérny Ig M sa v B lymfocytoch viaže na membránu, kde pôsobí ako receptor pre antigén. Sekretovaná forma Ig M vytvára pentamér (spojenie piatich Ig M, obr. 23), ktorý vďaka svojej veľkosti nemôže difundovať, a preto sa v medzibunkových tekutinách vyskytuje v nízkych koncentráciách. Udáva sa, že Ig M tvorí ako pentamér asi l0 % sérového imunoglobulínu. V rámci primárnej imunitnej odpovede (keď sa organizmus stretne s konkrétnym antigénom prvýkrát) sa Ig M produkujú ako prvé zo všetkých tried. Syntetizujú sa tiež ako prvé u novorodencov. Ig M sa v organizme tvorí najmä po imunizácii korpuskulárnym (nerozpustným) antigénom. Z tohto dôvodu sa významne uplatňuje v aglutinačných reakciách. Ig M je zo všetkých tried najúčinnejší vo väzbe vírusových častíc a aktivácii komplementu. Neprechádza placentou, ale môže byť transportovaný rovnakým mechanizmom ako Ig A prostredníctvom sekrečných epitelov.
- Ig A sa vyskytuje v dvoch formách: sérovej (v závislosti od živočíšneho druhu tvorí 1 - l0 % imunoglobulínov v krvnom sére) ako monomér, dimér, trimér alebo tetramér a slizničnej, ktorá je prevládajúcim imunoglobulínom v sekrétoch (sliny, slzy, hlien). Ig A je tiež hlavnou protilátkou v mlieku a kolostre, čím poskytuje ochranu mláďatám proti gastrointestinálnej infekcii patogénmi. Štruktúra slizničného Ig A je dimér alebo tetramér. Ig A funguje ako slabý opsonín, v imunokomplexe môže aktivovať alternatívnu cestu komplementu. Jeho hlavnou funkciou je ochrana slizníc, na ktoré sa viažu mikroorganizmy, zabraňuje ich adhézii a uľahčuje ich elimináciu.
- Ig D tvorí 0,2 % všetkých imunoglobulínov v krvnom sére. Pomerne vo vysokých koncentráciách sa nachádza v slzách. Produkujú ho tu spolu s Ig A, plazmocyty, ktoré pochádzajú z tonzíl a adenoidných vegetácií a migráciou cez cervikálne lymfatické uzliny sa dostávajú do slznej žľazy. Ig D má síce protilátkovú aktivitu, ale táto je v porovnaní s aktivitou ostatných tried imunoglobulínov nepatrná. Aktivuje aj komplement, ale len alternatívnou cestou. Jeho hlavná funkcia je receptorová - spoločne s Ig M je hlavným membránovým imunoglobulínom (receptor) v B lymfocytoch. Zatiaľ bola táto trieda opísaná len u človeka a myší.
- Ig E sa vyskytuje v sére zdravých jedincov v nepatrných koncentráciách (0,3 mg/ml). Zvýšené množstvá tohto imunoglobulínu sú identifikované u osôb s alergickými chorobami a u zvierat ťažko infikovaných parazitmi (Ig E sa vyskytuje napr. u ošípanej, psa, mačky). Z tohto dôvodu sa uplatňuje najmä v obranných reakciách proti mnohobunkovým parazitom na slizniciach a je hlavnou príčinou hypersenzitívnych (alergických) reakcií. Vyskytuje sa ako monomér. Ig E protilátky neprechádzajú placentou, nefixujú komplement, ale majú schopnosť viazať sa na viaceré efektorové bunky (eozinofilné granulocyty, makrofágy), ktoré sa zúčastňujú fagocytózy (obrany proti helmintom).
Molekulárna podstata tvorby protilátok
Odhaduje sa, že živočíšne druhy z triedy cicavce sú schopné vytvoriť až 1011 - 1012 rôznych protilátok. Protilátky, ako všetky proteíny, vznikajú procesom proteosyntézy. Vzhľadom na skutočnosť, že napr. genóm (súbor všetkých génov organizmu) človeka obsahuje „len“ desiatky tisíc génov, je zrejmé, že jednotlivé molekuly imunoglobulínov nemôžu byť kódované kompletnými génmi. Potomkovia nededia od rodičov celé gény pre polypeptidové reťazce imunoglobulínov, dedia iba segmenty týchto génov - subgény (génové segmenty). Subgény neobsahujú terminačný kodón, a preto nekódujú samostatne funkčný polypeptid. Až ich spojením počas diferenciácie B lymfocytu v kostnej dreni sa vytvoria príslušné gény pre molekuly L alebo H reťazca, pretože prvý segment obsahuje na 5' konci iniciačný kodón a posledný segment na 3' konci terminačný kodón. Transkripciou a transláciou obidvoch vytvorených génov vznikne jeden ľahký a jeden ťažký reťazec. Ich spojením disulfidickými väzbami vzniká funkčný imunoglobulín.
Subgény sa rozdeľujú do šiestich skupín:
- L-subgény kódujú signálny peptid (časť polypeptidového reťazca dôležitá pre transport molekuly v bunke) imunoglobulínového reťazca. Pred každým L-subgénom sa nachádza promótor (regulačná oblasť génu), ktorý je aktívny až po vytvorení celého génu.
- V-subgény kódujú variabilnú oblasť imunoglobulínového reťazca. Každému V-subgénu predchádza L-subgén, t. j. počet L- a V-subgénov je rovnaký.
- C-subgény kódujú konštantnú oblasť imunoglobulínového reťazca. Pred každým C- subgénom (okrem Cs) sa nachádza signálna sekvencia (s), ktorá je dôležitá pri vzniku imunoglobulínov tried Ig G, Ig A a Ig E.
- J-subgény kódujú 12 až 21 koncových aminokyselín variabilnej oblasti - tzv. spájajúcu oblasť.
- D-subgény sa podieľajú na kódovaní iba ťažkých (H) reťazcov imunoglobulínov, kde kódujú aminokyseliny tretieho hypervariabilného úseku.
- Subgény pre K reťazce sú u človeka lokalizované na chromozóme 2, subgény pre X reťazce sa nachádzajú na chromozóme 22 a subgény pre H reťazce na chromozóme 14.
Proces kombinovania subgénov sa uskutočňuje zvláštnym mechanizmom označovaným ako preskupovanie subgénov (somatická rekombinácia, VDJC rekombinácia). Výsledkom preskupenia subgénov je vznik jedného kompletného štruktúrneho génu pre L reťazec a jedného génu pre H reťazec. Preskupovanie, ktoré vedie k tvorbe génov pre ľahké reťazce sa uskutočňuje tak, že sa V-subgén spojí s J-subgénom za delécie (vylúčenia) celého úseku medzi nimi. Zachovaná zostáva len sekvencia medzi L a V a medzi V a C, ktorá v procese proteosyntézy podlieha transkripcii, posttranskripčným úpravám (splicing) a translácii. Výsledkom uvedeného procesu je jedna molekula ľahkého reťazca. Preskupovanie, ktoré vedie k tvorbe génov pre ťažké reťazce prebieha podobne: najskôr sa však D-subgén spojí s J-subgénom za delécie úseku medzi nimi a k tomu sa priradí V-subgén, pričom celý úsek medzi D-J a V-subgénom sa opäť vylúči. Po transkripcii a translácii vzniká jedna molekula ťažkého reťazca. Subgény VH, D, JH sa pri vzniku ťažkého reťazca preskupujú v kombinácii so subgénom C^, preto sa vytvorí ťažký reťazec imunoglobulínu Ig M. Súčasne so syntézou H reťazca pre Ig M sa tvorí aj H reťazec pre Ig D. Imunoglobulíny iných tried vznikajú tzv. prešmykom imunoglobulínových tried - deléciou úsekov medzi signálnymi sekvenciami (s), ktoré sa nachádzajú pred všetkými C-subgénmi (okrem Cs, ktorý signálnu sekvenciu nemá, preto H reťazec pre Ig D vzniká súčasne s H reťazcom pre Ig M). Vzhľadom na to, že na základe typu ťažkého reťazca (jeho konštantnej oblasti) zaraďujeme imunoglobulíny do jednej z piatich tried, prešmyk jednotlivých C-subgénov vedie k zmene triedy imunoglobulínov z Ig M na Ig A, Ig G alebo Ig E. Variabilná oblasť H a L reťazca pritom zostáva rovnaká, zmení sa len konštantná oblasť ťažkého reťazca.
Prešmyk imunoglobulínových tried prebieha až po skončení procesu preskupovania subgénov kódujúcich ťažký reťazec p. imunoglobulínu Ig M, a to až po stimulácii antigénom v sekundárnych lymfoidných orgánoch (nie počas vývoja B lymfocytu v kostnej dreni). Každý B lymfocyt obsahuje jeden štruktúrny gén pre ľahký reťazec a jeden pre ťažký reťazec. Okrem toho je expresia imunoglobulínových génov obmedzená vždy len na jednu alelu chromozómového páru, t. j. v príslušnom B lymfocyte sa expresiou vyjadrí len jedna alela pre H reťazec a jedna alela pre L reťazec. Tento jav sa nazýva alelová exklúzia. Pri vzniku imunoglobulínového reťazca sa uplatňuje tiež izotypická exklúzia: v každom B lymfocyte sa tvorí len jeden typ ľahkého reťazca - buď K alebo X.